ISGylation, the modification of protein targets by the interferon-stimulating gene protein 15 (ISG15), involves a cascade of enzymatic reactions utilizing the ubiquitin-activating enzymes E1, E2, and E3. These enzymes catalyze the conjugation of ISG15 to a lysine residue in target proteins.
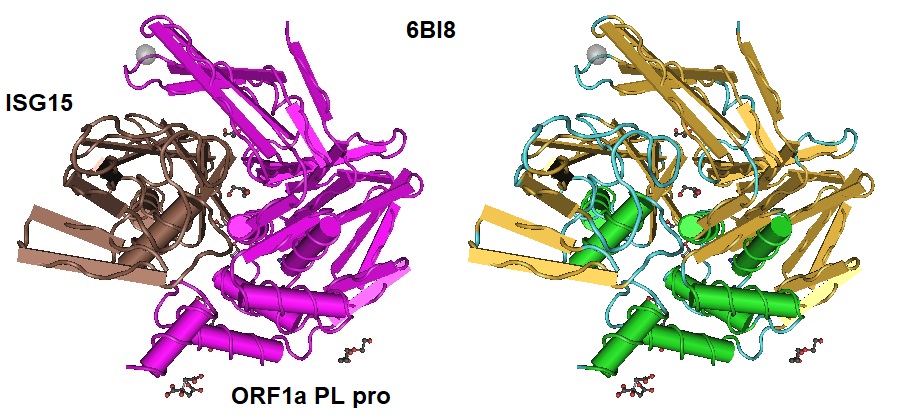
Figure 1: Human ISG15 in complex with MERS CoV papain-like protease. PDB ID 6BI8 (Clasman et al. 2018).
Sequences
>pdb|6BI8|C Chain C, Ubiquitin-like protein ISG15
MGWDLTVKMLAGNEFQVSLSSSMSVSELKAQITQKIGVHAFQQRLAVHPSGVALQDRVPLASQGLGPGSTVLLVVDKSDEPLSILVRNNKGRSSTYEVR
LTQTVAHLKQQVSGLEGVQDDLFWLTFEGKPLEDQLPLGEYGLKPLSTVFMNLRLRG
>pdb|6BI8|A Chain A, ORF1a
NDETKALKELYGPVDPTFLHRFYSLKAAVHGWKMVVCDKVRSLKLSDNNCYLNAVIMTLDLLKDIKFVIPALQHAFMKHKGGDSTDFIALIMAYGNCTFG
APDDASRLLHTVLAKAELCCSARMVWREWCNVCGIKDVVL QGLKACCYVGVQTVEDLRARMTYVCQCGGERHRQLVEHTTPWLLLSGTPNEKLVTTSTA
PDFVAFNVFQGIETAVGHYVHARLKGGLILKFDSGTVSKTSDWKCKVTDVLFPGQKYSSD

Figure 2: Structure of ISG15 at 2.5 Å resolution. PDB ID 1Z2M (Naraimhan et al. 2005).
As a response to viral or parasitic infections, the conjugation of the ISG15 protein to cellular targets induces signaling pathways that stimulate enhanced expression of temporally coordinated subsets of cell-type-specific proteins mediating anti-viral responses. Viral lipopolysaccharide and double-stranded RNA via interferon-α/β induce ISG15 conjugation.
Abnormal overexpression of ISG15 inhibits the replication of human immunodeficiency virus. Inhibition occurs via abolishing the nuclear processing of unspliced viral RNA precursors. This process is also known to block the replication of the influenza B virus. ISG15 is essential in innate immunity.
ISG15 is constitutively present in higher eukaryotes and has other functions not related to anti-viral defenses. The sequence of ISG15 terminates in the conserved LRLRGG motif.
The innate immune system of mammalian cells, including human cells, protects the cells against virus infection. RNA viruses activate innate immunity when pattern recognition receptors such as toll-like receptor-3, retinoic acid-inducible protein-I (RIG-I), and melanoma differentiation-associated gene 5 (MDA5) bind double-stranded viral RNAs. The result is the recruitment of adaptor proteins, toll-interleukin receptor domin-containing adapter-inducing interferon-B (TRIF), mitochondrial antiviral-signaling protein (MAVS), and assembly of signal complexes. The signal complexes that include E3 ubiquitin ligases (TRAF6, cIAP, TRAF3) and protein kinases (TAK1, IKKβ, TBK1, IKKε) activate transcription factors nuclear factor κB (NF-κB) and interferon-regulatory factor (IRF-3 and -7). This activated pathway initiates the production of interferon (IFN-α and IFN-β). The secreted interferon acts as an autocrine stimulant, now transcribing more than 300 interferon-stimulating genes (ISGs). This antiviral response is executed in infected and neighboring cells by cellular enzymes, transcription factors, cytokines, and chemokines.
The antiviral ubiquitin-like interferon-stimulating gene protein (ISG15) modifies cellular and viral proteins upon viral infection. The modification mechanism is like ubiquitination. The Ubiquitin-like protein plays a crucial role in the innate immune response to a viral infection either via its conjugation to a target protein (ISGylation) or via its action as a free or unconjugated protein. ISGylation starts a cascade of enzymatic reactions involving the ubiquitin-activating enzymes E1, E2, and E3. These enzymes catalyze the conjugation of ISG15 to a lysine residue in the target protein. Target proteins include interferon-induced protein with tetratricopeptide repeats 1 (IFIT1), interferon-induced GTP-binding protein MX1/MxA, protein phosphatase 1B (PPM1B), ubiquitin/ISG15-conjugating enzyme E2 L6 (UBE2L6), ubiquitin-like modifier-activating enzyme 7 (UBA7), and charged multivesicular body proteins 5, 2A, 4B and 6 (CHMP5, CHMP2A, CHMP4B, and CHMP6).
ISG15 can also ISGylate interferon-induced, double-stranded RNA-activated protein kinase (EIF2AK2/PFR), resulting in its activation. The activated pathway is known to have antiviral activity towards both DNA and RNA viruses, including influenza A, HIV-1, and Ebola virus.
Recently, new functions of ISGylation have emerged. ISGylation appears to be involved in other cellular processes such as DNA repair, autophagy, protein translation and exome secretion. Villarroya-Beltri proposed that under conditions of stress or infection, ISGylation acts as a defense mechanism. ISGylation is thought to inhibit normal protein translation by modifying protein kinase R (PKR), however, any newly synthesized proteins are tagged and marked as potentially dangerous. Following these events, the endosomal system is re-directed towards protein degradation at the lysosome. Hence, the cell's gates are locked and the spread of pathogens through exosomes is prevented.
Reference
Akutsu M, Ye Y, Virdee S, Chin JW, Komander D. Molecular basis for ubiquitin and ISG15 cross-reactivity in viral ovarian tumor domains. Proc Natl Acad Sci U S A. 2011 Feb 8;108(6):2228-33. [PMC]
Clasman, J.R., Everett, R.K., Mesecar, A.D.; Structural basis of Activity towards ISG15 for the Viral protease from MERS coronavirus. To be published.
ISG15 gene
Narasimhan J, Wang M, Fu Z, Klein JM, Haas AL, Kim JJ.; Crystal structure of the interferon-induced ubiquitin-like protein ISG15. J Biol Chem. 2005 Jul 22;280(29):27356-65. [JBC]
Carolina Villarroya-Beltri, Susana Guerra, Francisco Sánchez-Madrid; ISGylation – a key to lock the cell gates for preventing the spread of threats. Journal of Cell Science 2017 130: 2961-2969. [JCS]
---...---