In 2005, Pham & Sontheimer studied the molecular requirements for RNA-induced silencing (RNAi) complex (RISC) assembly in the Drosophila RNA interference pathway. The study used native gel electrophoresis to reveal components in the Drosophila RISC complex.
Component R1 forms when the Dicer-2/R2D2 heterodimer binds short interfering RNA (siRNA) duplexes. The heterodimer alone can initiate RISC assembly. Hence, other factors are dispensable for initiation. During assembly, R2 requires Argonaute 2 to convert into holo-RISC. This requirement reminds us of the RISC-loading complex, which also requires Argonaute 2 for assembly into RISC.
Pham & Sontheimer compared R2 to the RISC-loading complex and show that the two complexes are similar in their sensitivities to ATP and chemical modifications on siRNA duplexes. The researchers studied RISC formation requirements and showed that the siRNA 5'-termini are repeatedly monitored during RISC assembly. First by the Dcr-2/R2D2 heterodimer and again after R2 formation before siRNA unwinding. The 2'-position of the 5'-terminal nucleotide also affects RISC assembly because a siRNA strand bearing a 2'-deoxyribose at this position can inhibit the cognate strand from entering holo-RISC. The 2'-deoxyribose-modified strand has enhanced activity in the RNA interference pathway.
In 2007, Shah et al. observed that siRNA modified at the 5'-antisense phosphate can still cause RNAi, however not at the level affected by fully native siRNA. The researchers' result suggested an inherent tolerance of the RNAi machinery toward modification of the 5'-antisense phosphate.
In 2015, Prakash et al. reported that the electronic and spatial orientation of the 5′-phosphate analog is critical for single-stranded-siRNA activity. Chemically modified single-stranded-siRNA targeting human ApoC III mRNA showed enhanced potency for inhibiting ApoC III mRNA and protein in transgenic mice. The modified siRNAs were able to reduce the triglyceride and LDL cholesterol in transgenic mice.
In 2017, Haraszti et al. showed that 5΄-Vinylphosphonate modification of siRNAs protects them from phosphatases, improves silencing activity, and confers novel properties to siRNAs.
5΄-vinylphosphonate (i) increases siRNA accumulation in tissues, (ii) extends the duration of silencing in multiple organs, and (iii) protects siRNAs from 5΄-to-3΄ exonucleases.
Because 5΄-phosphate is required for loading into RNA-induced silencing complex, a 5΄-phosphate on a fully modified siRNA guide strand is expected to be beneficial. The synthetic phosphorylation of fully modified cholesterol-conjugated siRNAs increases their potency and efficacy in vitro. Unfortunately, when delivered systemically to mice, the 5΄-phosphate is removed within 2 hours.
The 5΄-phosphate mimic 5΄-(E)-vinylphosphonate stabilizes the 5΄-end of the guide strand by protecting it from phosphatases and 5΄-to-3΄-exonucleases. The improved stability significantly enhances the efficacy of cholesterol-conjugated siRNAs and the duration of silencing in vivo. 5΄-(E)-vinylphosphonate stabilizes 5΄-phosphate and enables systemic delivery to and silencing in the kidney and heart.
Table 1: RNAi modifications of the 5’-Phosphate and their Effect.
|
Modification
|
Structure
|
ΔTm duplex per modification
|
Impact on the efficiency of RNAi
|
Others
|
|
5′-C-methyl
(S-isomer)
|
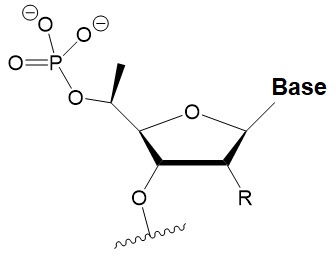
|
−3.2°C
|
One (S) 5′C-methyl at the 5′-end of the antisense strand is tolerated by RNAi.
|
The introduction of two amide linkers from the 3′-ends of the duplex increases the nuclease resistance of siRNA in serum.
|
|
5′-(E)-vinyl-phosphonate
|
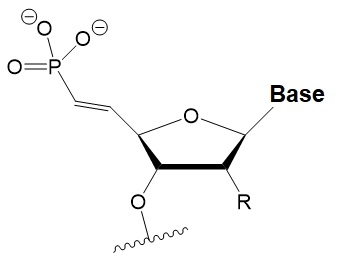
|
No data.
|
5′(E)-vinyl-phosphonate improves siRNA interaction efficiency with Ago2 by 2-fold.
Does not change the biological activity of siRNA in vitro.
|
Stabilizes 5′-phosphate. Protects from the action of phosphatases and exonucleases.
Improves the pharmacokinetics.
Enables systemic delivery to and silencing in kidney and heart.
|
|
5′-methylene-phosphonate
|

|
No data.
|
5′-methylene-phosphonate at the 5′-end of the antisense strand reduces the biological activity of siRNA by ~10-fold.
|
No data.
|
Reference
Chernikov IV, Vlassov VV, Chernolovskaya EL. Current Development of siRNA Bioconjugates: From Research to the Clinic. Front Pharmacol. 2019 Apr 26;10:444. [PMC]
Haraszti RA, Roux L, Coles AH, Turanov AA, Alterman JF, Echeverria D, Godinho BMDC, Aronin N, Khvorova A. 5΄-Vinylphosphonate improves tissue accumulation and efficacy of conjugated siRNAs in vivo. Nucleic Acids Res. 2017 Jul 27;45(13):7581-7592.[PMC]
Pham JW, Sontheimer EJ. Molecular requirements for RNA-induced silencing complex assembly in the Drosophila RNA interference pathway. J Biol Chem. 2005 Nov 25;280(47):39278-83. [PMC]
Shah S, Friedman SH. Tolerance of RNA interference toward modifications of the 5' antisense phosphate of small interfering RNA. Oligonucleotides. 2007 Spring;17(1):35-43. [PubMed]
Thazha P. Prakash, Walt F. Lima, Heather M. Murray, Wenyu Li, Garth A. Kinberger, Alfred E. Chappell, Hans Gaus, Punit P. Seth, Balkrishen Bhat, Stanley T. Crooke, Eric E. Swayze; Identification of metabolically stable 5′-phosphate analogs that support single-stranded siRNA activity. Nucleic Acids Research, Volume 43, Issue 6, 31 March 2015, Pages 2993–3011. [PMC]
---...---